It is possible that the Makganyene glaciation played a role in the evolution
of eukaryotes and the Marinoan glaciation in the achievement of multicellularity
in (microscopic) animals. However, the fossil record provides no support
for close correlations in time and causal links are entirely speculative.
In the case of the
Makganyene
glaciation, we are talking about the origin
and
adaptation of
eukaryotic cells.
With the Sturtian and Marinoan glaciations, the origin of macroscopic (visible
to the naked eye) multicellular animals. The oldest
body fossils believed to represent a eukaryotic organism, mainly on account
of their size, is the cm-scale coiled form
Grypania,
which occurs in strata ~1.89 Ga in age in northern Michigan (USA). They
are >300 Myr (million
years) younger than the Makganyene glaciation ~2.22 Ga. Biomarkers (fossil
organic molecules) attributed to eukaryotes are reported from strata nearly
2.7 Ga in age in Western Australia, but the phylogenetic specificity of
the molecules has been questioned. There are far too few data to gain traction
on a possible link between the Makganyene snowball earth and the origin
of eukaryotic organisms.
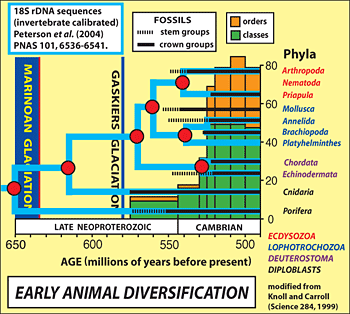
As for early animals, it is useful to couch the question in terms of an
evolutionary "tree", given the new light that molecular data
are shedding on early animal phylogeny. Multicellularity in animals (multicellularity
evolved much earlier in
algae) is a trait common to
calcisponges and
eumetazoa,
and presumably their last common ancestor. The genetic "tool-kit" for
multicellularity (i.e., genes for inter-cell adhesion, signalling and differentiation)
already exist in
choanoflagellates, unicellular (occasionally colonial)
protists that are "rooted" more deeply in the animal tree than
the calcisponge-eumatzoa divergence. Molecular-clock analysis using a concatenation
of different amino acid sequences, calibrated by known invertebrate branching-points
across the Phanerozoic, places the calcisponge-eumetazoa divergence at roughly
650 Ma according to
Kevin Peterson and coworkers at Dartmouth College (New
Hampshire, USA). The next major divergence was the branching of the stem
eumetazoa into
cnidarians and
bilaterians. The stem metazoa had evolved
from eating bacteria to eating eukarya, a huge leap in the size of prey.
The same molecular clock places the cnidaria-bilateria divergence at roughly
615 Ma, which is approximately coincident with an abrupt increase in the
size, diversity, spiny morphology and turnover rate of fossil eukaryotic
plankton, suggested by Peterson to be a response to predation. The absence
of recognizeable body fossils or trace fossils at this time implies that
animals were still microscopic.
The bilateria next divided into
deuterostomes (echinoderms and hemichordates,
which later gave rise to vertebrates including our own species) and
protostomes.
The last common ancestor to all bilateria (LCB) had a body plan that was
differentated along a central axis and also top-to-bottom (dorsal-ventral).
Molecular-clock analysis places it around 570 Ma, which is close to the
first appearance of
Ediacara, abundant soft-bodied macrofossils (up to 2
m in length), commonly frond-like, whose modular construction is unlike
any extant animal. There is increasing doubt that they were animals at all,
a view championed by
Dolf Seilacher at the University of Tübingen (Germany)
originally considered heretical. The last common protostome (LCP) branched
into the
ecdysozoa (animals that moult their exoskeletons, including arthropods
and priapulids) and the
lophotrochozoa (bivalves, annelids, platyhelminthes,
etc.). Peterson's molecular-clock analysis places the LCP at roughly
550 Ma, which is not too far off the age of the oldest fossil lophotrochozoan,
the 555-Ma stem-group mollusc
Kimberella from
Arctic Russia, described by
Mikhail Fedonkin of the Paleontological Institute in Moscow (Russia). Evidence
for locomotion on the sea-floor by the use of appendages is not seen in
the trace-fossil record until the base of the
Cambrian at 542 Ma, and the
canonical Cambrian "
explosion" of bilaterian skeletal diversity
was delayed until 526-520 Ma (Tommotian and Atdabanian Stages of the Early
Cambrian).
From the foregoing, it would seem that the achievement of multicellularity in
(microscopic) animals would be the evolutionary step most closely associated
in time with the 635-Ma (Marinoan) snowball earth. However, there is as yet no
empirical support for this in the fossil record.

